
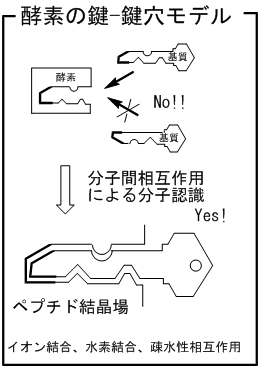
 酵素は鍵−鍵穴モデルで示される特異な分子認識
酵素は鍵−鍵穴モデルで示される特異な分子認識
場を発現します。酵素を構成するタンパク質は,ア
ミノ酸がペプチド結合によりつながり一次構造を形
成し,それらが二次構造・三次構造と組みあがるこ
とで基質を認識する鍵穴を形成します。そして鍵穴
にマッチした鍵となる基質のみを選択的に取り込ん
で反応します。例えば,セリンプロテアーゼのキモト
リプシンは245のアミノ酸から構成させています。
それでは,このようなタンパク質の機能をモデル
化するのにどれくらいの長さのぺプチドが必要で
しょうか?
たった二分子のアミノ酸からなるジペプチドも
結晶として配列することにより,酵素と同様の分
子間相互作用により,基質を立体選択的に取り込
みます。このような有機ホスト分子がゲスト分子
を取り込んだ化合物を包接化合物といいます。
・ジペプチド包接結晶による分子認識
ジペプチドは双性イオンのため、その静電相互作用と水素結合によってシート構造を形成します。そのシートが積層する際にアミノ酸の側鎖が壁となりシート間に分子を認識するキラルな空孔を形成します。このような最も単純な酵素モデルとしてジペプチド分子を用いる分子認識について研究しています。
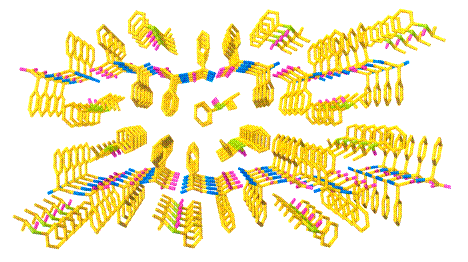
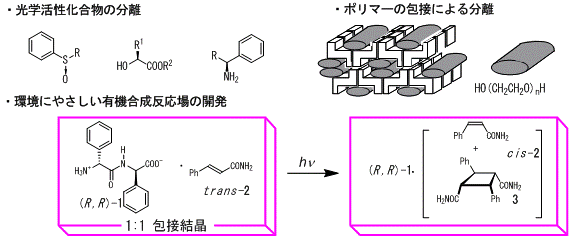
・ジペプチド包接結晶の機能と応用
ジペプチド結晶の包接現象を利用して,さまざまな機能について研究しています。ジペプチドの形成する空孔はキラルであるため光学活性化合物の光学分割が可能です。またチャネル状の空孔を利用した高分子の分離も可能です。また,取り込み場での光反応も可能であり,酵素モデルの固体触媒を目指して研究しています。
このように生体構成分子であるアミノ酸(ペプチド類)の結晶工学の観点から、分子配列の制御により、新しい人工酵素の構築とその分子間相互作用を解明しています。